|
|

|
Plankton - Eine Übersicht

|
| Fundort |
Was hier geleistet werden soll ist eine kleine Übersicht über die wichtigsten Gruppen von Kleinstlebewesen, die sich vor allem unter dem Mikroskop bei geringer Vergrößerung beobachten lassen.
Das sind also im Wesentlichen große (eukariotische) Einzeller und kleine Vielzeller bis zu einer Größe von etwa einem Millimeter.
Im Grunde also das, was man in Mitteleuropa im Sommer in einer Pfütze, einem Teich oder auch in einem Heuaufguss mit dem Mikroskop entdecken kann - und natürlich auch in einem Aquarium.
Die Bezeichnung Plankton ist daher hier ziemlich ungenau verwendet - nennen wir das mal umgangssprachlich.
Genauer wäre der weniger bequeme Begriff Mikroorganismen.
Näheres wie Plankton streng definiert ist steht weiter unten.
Bakterien sollen hier nicht betrachtet werden, denn sie sind im Lichtmikroskop wenig reizvolle Objekte.
Von den korkenzieherartig geformten Bakterien abgesehen (die sich mitunter sehr schnell bewegen) erscheinen sie meist punkt- komma- oder stäbchenförmig.
Ihre genauere Beschaffenheit lässt sich nicht erkennen.
(Ihre Bestimmung im Labor erfolgt nicht im Lichtmikroskop - untersucht wird zunächst Wuchs, Form und Farbe einer Kolonie in der Petrischale.)
Dieses Thema ist so umfangreich, dass eine einzelne Internetseite auch dem Anspruch, nur eine kleine Übersicht abliefern zu wollen kaum gerecht werden kann.
Wer sich dafür interessiert dem sei die unten angeführte Literatur empfohlen.
Was ist Plankton ? - Definitionen und Abgrenzungen
Wie erwähnt gibt es eine strenge Definition von Plankton.
Sie legt solche Organismen als Plankton fest, die vorwiegend im Wasser treiben, deren Bewegungsrichtung also von der Strömung vorgegeben ist und der sie aus einenem Antrieb kaum oder keinen Widerstand leisten können.
Das müssen nicht unbedingt Mikroorganismen sein, auch für etliche Fischlarven oder sogar manche Staatsquallen trifft das zu.
Allen Versuchen, die Welt der Mikroorganismen einzuteilen muss eine grundsätzliche Sache vorangestellt werden:
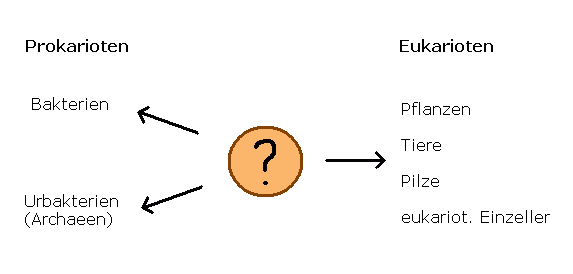
Alles Leben dieser Welt lässt sich einteilen in zwei Gruppen:
Zum einen solche Lebewesen, deren Zellen einen Zellkern aufweisen, der das Erbgut in sich birgt und es mit einer Membran von Rest der Zelle trennt.
Dazu zählen alle höhrenen Lebewesen wie Pflanzen, Tiere und Pilze ("eukariotisch").
Zum anderen sind das Organismen, die keinen Zellkern besitzen ("prokariotisch").
Sie sind stets einzellig und bilden die Gruppe der Bakterien und Urbakterien.
Eine Fülle von weiteren fundamentalen Unterschieden trennt die beiden Gruppen der eukariotischen und prokariotischen Lebewesen, auf sie soll hier nicht näher eingegangen werden.
(Ein Lehrbuch der Mikrobiologie verglich die prokaryotische Zelle mit einem Schlauchboot, während die eukariotische Zelle hinsichtlich Größe und Organisationsgrad eher einem Tanker gleiche.)
Die Bezeichnung Mikroorganismen umfasst folgende Gruppen von einzelligen oder kleinen, mehrzelligen Lebewesen:
-
Urbakterien: Leben z.T. in großer Hitze, Druck oder Säure. Am vulkanischen Meeresgrund.
-
Bakterien: Enorm umfangreiche Gruppe, von Milchsäurebakterien bis Krankheitserregern.
-
Mikroalgen: Da es auch große Algen gibt (Seetang) ist eine Abgrenzung erforderlich. Mikroalgen sind meist einzellig oder in kleinen Verbänden organisiert.
-
Pilze: Einzellige Pilze wie Backhefe
-
Protozoen: Eukariotische Tiere, die als Einzeller oder in kleinen Verbänden leben. Als Tiere sollten sie beweglich sein und sich nicht durch Photosynthese oder Vorgänge aus der anorganischen Chemie ernähren.
-
Viren: Da sie keinen eigenen Stoffwechsel aufweisen und sich nur über Wirte vermehren können, ist ihr Status als echte Lebewesen (zu recht) umstritten.
Der Begriff Protisten ist ein Überbegriff und bezeichnet die eukariotisachen Mikroorganismen, also die (pflanzlichen) Mikroalgen, Pilze und die (tierischen) Protozoen.
Protisten leben für gewöhnlich im Wasser; sie treten global auf und es ist wohl noch kein Ort gefunden worden, von der Tiefsee bis zu den Gebirgsgipfeln, an dem sie nicht auftreten.
Ihre oft immensen Individuenzahlen machen sie zu einem der heimlichen Herrscher des Ökosystems Erde.
Gerade kleine Lebensräume wie wassergefüllte Baumstümpfe oder der Flüssigkeitsfilm auf Steinen in direkter Nähe zu einem Wasserfall werden von ihnen besiedelt.
An ihnen ist das urtümliche Leben sichtbar, aus dem im Laufe der Evolutin durch Koloniebildung komplexe Vielzeller entstanden sind.
Während kleine aber vielzellige Mikroorganismen noch Zellverbände hervorbringen können, die eine bestimmte Aufgabe bewältigen, (z.B. der Kaumagen der Rädertiere) müssen Einzeller entsprechende Funktionseinheiten innerhalb einer Zelle erstellen.
Analog zu den vielzelligen Organen der Vielzeller (Magen, Niere) bilden sie Organellen wie Wimpern oder den Zellmund aus.
Protisten verfügen meist über ein enormes Vermehrungspotential.
Meist folgen viele Generationen von nicht-sexueller Reproduktion aufeinander (Teilung bei Einzellern, Parthenogegese bei kleinen Vielzellern);
in Mangelzeiten oder auch spontan wird zur sexuellen Reproduktion übergegangen, aus der Dauerstadien hervorgehen können.
Sie werden mit der Strömung, diversen Trägern wie Wasservögeln oder auch mit dem Wind verfrachtet.
Vorgänge und Organellen, die bei Protisten gut zu beobachten sind spielen auch bei Zellen innerhalb von Vielzellern noch mitunter eine große Rolle;
Flimmerhärchen auf der Lungenoberfläche, die der Selbstreinigung dienen gehen auf die Fortbewegungsgeißeln von frei beweglichen Einzellern zurück.
Wasser ist etwa 800 Mal dichter als Luft - ein gehöriger Unterschied.
Gerade bei kleinen Abmessungen wirkt das als ob die Organismen mehr durch ein Gelee als durch eine Flüssigkeit schwimmen.
Außerdem sind dadurch und auch bedingt durch die Kleinheit der Organismen Formen und Baukonzepte möglich, die an Land oder bei größeren Abmessungen unweigerlich unter dem eigenen Gewicht zusammenbrechen würden.
Ebenso tritt der Organismus direkt in Kontakt mit weiteren chemischen Umgebungsfaktoren.
Während es an der Luft im wesentlichen nur Temperaturänderungen gibt - auch der Sauerstoffgehalt ist konstant - schwanken im Wasser pH-Wert und eine Fülle von gelösten Stoffen in ihrer Konzentration.
Und selbst der Sauerstoffgehalt ist variabel und kann unter ungünstigen Umständen gegen Null tendieren - ein Umstand, dem mit einer desto heftigeren Vermehrung zu günstigen Zeiten Rechnung getragen wird.
Das Problem der Systematik
In dem Bestreben, die Fülle der Lebensformen in Gruppen zu ordnen und diese in Übergruppen zusammenzufassen hat sich im Laufe der Jahrzehnte und Jahrhunderte ein fein verästelter Stammbaum des Lebens gebildet, der mehrfach umgebaut wurde und heute vor einem seiner größten Umbauten steht.
Zwar lässt sich eine Art als Fortpflanzungsgemeinschaft noch einigermaßen klar definieren, alles was darüber hinausgeht, stellt stets Meinung dar, im Zweifelsfalle eben die Mehrheitsmeinung der Lehrstuhlinhaber.
Darwin selbst erkannte, dass aus seiner Evolutionstheorie folge, dass eine Systematik in der Biologie die Abstammungsgeschichte widerspiegeln müsse,
dass also zwei Gruppen, die ähnlich sind weil sie einen gemeinsamen Vorfahren haben auch in einer gemeinsamen Übergruppe zusammengefasst werden können (natürliche Systematik).
Nun hat man sich seit Erfindung des Mikroskops sehr auf das Aussehen eines Lebewesens konzentriert und daher ähnlich aussehenden Lebewesen einen gemeinsamen Vorfahren unterstellt.
Das hat sich gerade bei Mikroorganismen wiederholt als Fehler erwiesen.
Auch hat der technische Fortschritt mit neuen, besseren Auflösungen in der Mikroskoptechnik und neuen Analyseverfahren seinen Teil dazu beigetragen,
dass Inhaber von Lehrstühlen immer wieder in den Genuss kamen, die Meinungen ihrer Vorgänger korrigieren zu dürfen.
Hier ein minimalistischer Abriß der obersten Ebene.
Es zeigt die Einteilung aller Organismen in die Reiche und ihre Veränderung im Laufe der Zeit.
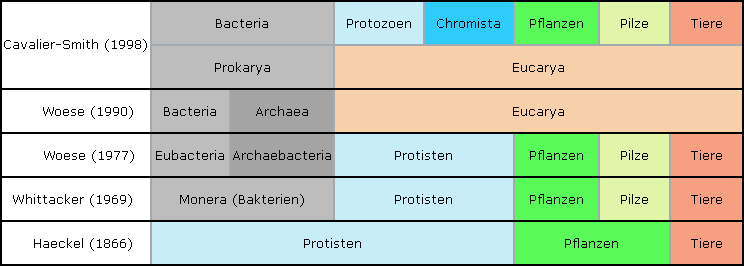
War Linne (1707 - 1778), einer der Väter der modernen Systematik noch von der Unveränderlichkeit der Arten überzeugt, so gelang es Ernst Haeckel (1834 - 1919), Darwins Theorie in Deutschland seinerseits bekannt zu machen.
Haeckel, eine hochinteressante, schillernde und respektable Persönlichkeit, teilte die Natur augenfällig ein in Pflanzen, Tiere und .. naja, alles was sehr klein ist.. eben Protisten.
Whittaker (1969) war bereits klar, das Pilze sich so sehr von Pflanzen unterschieden, dass sie ein eigenes Reich erforderten.
Ebenso waren hatten Bakterien aufgrund ihres prokariotischen, einfachen Zellaufbaus einen eigenen Platz gefunden.
Woese unterschied nun bei den Prokarioten zwischen Bakterien und Urbakterien, die vermutlich aufgrund ihrer teils extremen Lebensbedingungen vorher noch gar nicht entdeckt waren.
Auf der obersten Skala hatten nun sechs Haupt-Gruppen des Lebens Platz genommen.
1990 vereinfachte man das System und erstmals ist zu sehen, dass nicht Merkmale, die in einem Mikroskop zu sehen sind das entscheidende Kriterium darstellen, sondern der intrazelluläre Aufbau (bzw die molekulare Struktur der rRNA).
Woese hält aber die Unterschiede zwischen Bakterien und Urbakterien für so gravierend, daß er beiden ein eigenes Stellung zukommen läßt.
Die Einteilung erfolgt nun an oberster Stelle nicht mehr in Reiche sondern in Domänen.
Cavalier-Smith schließlich fasst die prokariotischen Bakterien und Urbakterien wieder zusammen und stellt sie als eine Domäne den eukariotischen Lebewesen gegenüber.
Bei den Eukarioten unterteilt er nun die Protisten in die (eher pflanzlichen) Chromista und die (eher tierischen) Protozoa ein.
Die eigentliche Revolution steht aber nun bevor:
Sina M. Adl (ein Lehrstuhlinhaber aus Kanada) et al. teilen 2005 das Leben streng nach phylogenetischen Merkmalen ein, das heißt die genetische Verwandschaft ist entscheidend.
Das wurde möglich nachdem große Mengen an Genen innerhalb überschaubarer Zeiträume sequenziert werden konnten.
Große Übereinstimmung im sequenzierten Genmaterial beweist nahe Verwandschaft.
Gleichzeitig wendet man sich ab von einem System, das auf historisch gewachsenen Unterscheidungen basiert.
Die Gruppe der Einzeller ist keineswegs eine Gruppe von ähnlichen Organismen, die - wie es die Einteilung voraussetzt - untereinander enger verwandt sind als zu jedem Vielzeller.
Die Einteilung in Protozoen und Nichtprotozoen war immer schon problematisch, da sich Vielzeller stets aus einer bestimmten Gruppe Einzeller entwickelt haben.
Ein wenig gleicht der Sachverhalt auch der Stellung der Vögel im Stammbaum des Lebens, die traditionell als eine der sechs Gruppen der Wirbeltiere auf gleicher Ebene neben den Reptilien stehen,
während man längt weiß dass sich Vögel aus Reptilien entwickelt haben und daher eine Teilgruppe von ihnen darstellen müßten.
Um das zu korrigieren hat man die Vielzeller bis hin zu Landpflanzen und Säugetieren dorthin eingeordnet wohin sie logisch auch gehören: Als Untergruppen derjenigen Einzeller, aus denen sie sich jeweils entwickelt haben.
Für die Pflanzen ist das womöglich eine kleine einzellige Grünalge namens Mesostigma,
vielzellige Tiere werden eingeordnet in die Gruppe der Opisthokonta, die vor allem kleine Einzeller umfaßt, die mit einer einzelnen Geißel am hinteren Ende ausgestattet sind.
Die Gesamtheit aller eukariotischen Lebewesen wird nun unterteilt in folgende Gruppen:
-
Amoebozoa:
Einzeller mit formveränderlicher (amöboider) Gestalt.
- Myxomyceten (Schleimpilze):
Hochinteressante Lebensform. Anfänglich einzellig lebend, dann sich zu einem Plasmodium vereinend, schließlich Bildung eines Fruchtkörpers.
- Thecamoeben (Schalenamöben):
Schutzhülle aus Chitin, teils mit Kieselsäure.
- Nacktamöben:
Das bekannte Wechseltierchen.
-
Opisthokonta:
- Kragengeißeltierchen (Choanoflagellaten):
Fadenförmige Zellfortsätzen bilden kreisförmig angeordnet eine Wulst, aus dessen Mitte eine einzelne Geißel entspringt.
- Pilze
- Metazoa:
Tierische Vielzeller, von Regenwurm bis Schäferhund.
- Mesomycetozoa:
Einzeller, meist Fischparasiten.
-
Rhizaria:
- Foraminiferen (Kammerlinge):
Meist im Meer. Einzeller, die in einem meist mehrkammerigen Gehäuse leben. Durch feine Poren treten dünne, untereinander vernetzte Scheinfüßchen aus. Formenreiche Gehäusestruktur.
- Strahlentierchen (Radiolaria):
Innenskelett aus Opal (Siliziumdioxid), das in Form einer durchlöcherten Kugel die Zelloberfläche abstützt. Von der Kugeloberfläche radial abstehende Stützen stabilisieren Fortsätze an der Zelloberfläche. Immer marin.
- Cercozoa
-
Archaeplastida:
- Glaucocystaceae:
Vertritt eine der drei Hauptlinien bei der Endosymbiontenentwicklung
- Rotalgen:
Rötlich gefärbt. Stets vielzellig, pflanzenartig. Dreigliedriger Generationenwechsel. Kein Chlorophyll b. Beliebte Sushi-Beilage.
- Chloroplastida (Grünalgen im weitesten Sinne. Gattungsbasis: Mesostigma):
- Grünalgen (Chlorophyta)
- Charophyta
- Schmuckalgen
- Streptophytina
Chromalveolata:
- Cryptophyceae:
Einzellige Mikro-Algen, zwei Geißeln, zwei Zellkerne. Nur teilweise mit der Fähigkeit zur Photosynthese. Typischerweise nicht als Tier oder Pflanze klassifizierbar.
- Kalkalgen (Haptophyta):
Planktisch lebende, photosynthetisch aktive Einzeller; zwei gleich lange Geißeln je Zelle. Manche Arten mit kalkigen Schuppen auf der Oberfläche.
- Stramenopile (= Chromista = Heterokonta):
Zwei unterschiedliche Geißeln. Meist photosynthetisch lebende Einzeller. Eustigmatales (z.B. Nannochloropsis), Netzschleimpilze, Goldbraune Algen, Gelbgrüne Algen, Braunalgen (immer mehrzellig, z.B Seetang), Kieselalgen u.a.
- Alveolata:
Apicomplexa (parasitische Einzeller, z.B. Malariaerreger), Dinoflagellaten (formenreiche Gruppe mit zwei senkrecht aufeinander stehenden Geißeln), Wimpertierchen (Ciliophora).
Excavata:
Charakteristische tiefe Mundöffnung.
- Fornicata
- Parabasalia
- Heterolobosea:
Amöboid lebende Einzeller. Fließamöben (Vahlkampfia), zelluläre Schleimpilze u.a.
- Euglenozoa:
Als einziger der Excavata betreiben manche Euglenozoa Photosynthese. Eine bekanntere Gattung hier: Augentierchen (Euglena sp.)
Die Gruppe der Protosten oder Protozoen hat mit diesem System aufgehört zu existieren.
Woraus sich die ersten Einzeller entwickelten ist nach wie vor unklar und wird wohl nie sicher nachzuvollziehen sein.
Auch wie Frage wie wahrscheinlich es ist dass aus unbelebter Materie unter geeigneten Bedingungen Leben entsteht wurde lange für unbeantwortbar gehalten.
Immerhin wäre für eine Statistik eine Vielzahl von Planeten nötig, die allesamt gleiche und geeignete Lebensbedingungen aufweisen.
Die Zahl derjenigen Planeten, die dann tatsächlich Leben hervorbringen, wäre dann ein Maß dafür, ob die Entstehung von Leben eine außergewöhnliche Ausnahme ist oder ob es dafür eine nennenswerte Wahrscheinlichkeit gibt.
Nun, man hat nur die Erde und kann aus einem einmaligen Ereignis keine Wahrscheinlichkeit ableiten.
Andererseits ist der Lauf der Evolution auch in den ersten Anfängen eine Aneinanderreihung von vielen Einzelereignissen, so dass man aus dem zeitlichen Ablauf sehr wohl auf eine Wahrscheinlichkeit schließen kann.
Dabei bedeutet eine schnell einsetzendes Ereignis eine hohe Wahrscheinlichkeit dafür dass es passiert.
Im Falle der Erde sind die allerersten einfachsten Lebensformen sehr früh entstanden, in geologischen Maßstab direkt nach ihrer Geburt, zu einer Zeit, als noch sehr viel häufiger Asteroiden auf den Planeten stürzten.
Einschläge von jenem Kaliber, der später die Dinosaurier ausrottete waren dabei wohl mehr die Regel als die Ausnahme.
Heute vermutet man daher das Leben sehr wahrscheinlich immer dann entsteht wenn geeignete Bedingungen herrschen,
vermutlich ist es auf der Erde sehr schnell und sogar mehrfach entstanden, mitunter parallel an verschiedenen Orten,
wurde ausgelöscht von einer kosmischen Katastrophe eines Einschlags und entstand erneut woanders, bis das kosmische Bombardement im frühen Sonnensystem weitgehend versiegte.
Die gleiche Überlegung aber muss auch in Rechung stellen, dass auf der Erde während des Großteils ihrer bisherigen Existenz nur primitive Einzeller lebten.
Von den - sagen wir - drei Milliarden Jahren des Lebens waren zwei den Einzellern vorbehalten, komplexe Vielzeller entstanden in größerer Zahl erst vor 600 Mio Jahren (das ist ein Fünftel der Zeit).
Und der Mensch ließ sehr lange auf sich warten.
Überträgt man diese Zeitspannen wieder in Wahrscheinlichkeit, so sind Vielzeller einigermaßen unwahrscheinlich, eine Zivilisation sogar sehr unwahrscheinlich. (Wobei die Durchschnittslebensspanne einer Zivilisation schwer abzuschätzen ist)
Ganz klar: Diese Wahrscheinlichkeiten sind hoch interessant um sie auf Exoplaneten anzuwenden - wie wahrscheinlich ist Leben in der Galaxis, wie viel werden wir finden?
Trifft diese Überlegung zu, so ist es durchaus möglich, Einzeller auf benachbarten Planeten zu finden.
Ich für meinen Teil bin zuversichtlich dass in den nächsten 30 Jahren extraterrestrischen Chlorophyll nachgewiesen wird.
Höher entwickeltes Leben oder gar eine Zivilisation sind jedoch sehr unwahrscheinilich.
Diese Erkenntnis brachte man mit dem geflügelten Wort auf den Punkt, nach dem in der Galaxis "grüner Schleim häufig, schöne Frauen aber selten" zu finden sind.
×
Ein weiteres Mal sieht der Mensch hier sich seiner Krone beraubt, die er sich selbst verliehen hat:
Ordnet man ihn wie alle anderen tierischen Vielzeller ein in die entsprechende Gruppe von Einzellern, aus denen er durch Koloniebildung entstanden ist,
so erscheint dieser Körper nur als Erfüllingsgehilfe der Eizelle, aus der er entstanden und deren Genschatz er weitergeben und so erhalten muss.
Man ist versucht, die einhundert Billionen Zellen als Arbeiterinnen zu sehen, die für eine einzelne Königin, die Eizelle, leben und sterben.
Soll das auch für das Gehirn gelten, dem Ort wo der Mathematik nachgespürt und Musik verstanden wird ?
Es ist eines der bittersten und gemeinsten Effekte der Evolution, dass der menschlichen Art gerade so viel Verstand erwuchs um zu erkennen dass der einzelne Mensch sterben muss, aber eben nicht genug um mit dieser Erkenntnis geistig fertig zu werden.
Aus dieser Lücke erwachsen Kulte und Religionen. Und vielleicht aus die Ruhelosigkeit des Forschergeists.
Nur, Kulte und Religionen, Mathematik und Musik haben mit der Evolution und der Keimbahn nichts oder fast nichts mehr zu tun.
Die Keimbahn ist der Mutterboden, die Ursuppe geworden, aus der der menschliche Geist auskeimt, ein Ereignis das genaus so folgenreich sein dürfte wie die Entstehung des Lebens selbst.
Als lebender Biologie-Forscher beginnt er das Leben selbst zu begreifen.
Selbstbewusst und verantworrtungsvoll soll er seine Krone tragen, seine Mathematik und Musik, von Generation zu Generation.
×
Pflanzlich oder Tierisch - Endosymbionten
Im Unterschied zu bakteriellen Zellen (Prokaryoten) besitzen die höher entwickelten Zellen (Eukatioten) Bauteile, die nicht von der Zelle selbt hergestellt werden können.
Es handelt sich dabei zum einen um die Mitochondrien, die in allen Zellen vorkommen und die Funktion eines Kraftwerkes übernehmen, welches den Energieträger ATP erzeugt;
zum anderen um die Plastiden, die in pflanzlichen Zellen die photosysthetisch aktiven Farbstoffe, v.a. das Chlorophyll enthalten.
Bei einer Zellteilung der eukariotischen Zelle muss die Mutterzelle zwangsläufig ihres Bestand an Mitochondrien oder Plastiden aufteilen und an beide Tochterzellen weitergeben.
Dort vermehren sich die Mitochondrien und die Plastide und erreichen die normale Anzahl in einer Zelle.
Beim Menschen beispielsweise vererbt sich daher die DNS der Mitochondrien immer von der Mutter zu den Nachkommen, da die Mitochondrien der Spermazelle nicht in die Eizelle miteindringen bzw. dort aufgelöst werden.
Das Genographic Project macht sich diesen Umstand zunutze und untersucht seit 2005 die Verwandschaftsbeziehungen von Volksgruppen, Stämmen und geht der Frage nach, wie groß die Urpopulation des Homo sapiens war ("Mitochondriale Eva", Ur-Eva).
Des weiteren ähnelt die DNS der Mitochondrien und der Plastiden eher der von Bakterien;
sie liegen nicht als Faden sondern als geschlossener Ring vor, sind anders aufgewickelt (keine Histone),
enthalten kaum repetitive DNA (Junk-DNA) und einige Merkmale, die beim Umsetzen der DNA in Proteine Signalcharakter haben, entsprechen eher den entsprechenden Signalen der Bakterien.
Beide Organellen sind von einer Doppelmembran umgeben; die innere änelt einer Bakterienmembran, die äußere ähnelt der Außenmembran der Eukarioten.
Der Mechanismus, der die DNA in lebende Proteine umsetzt (die Ribosomen) ähneln so sehr denen der Bakterien, dass sie ebenfalls auf Antibiotika reagieren, die sonst nur Bakterien schädigen.
Alles in allem also eine durchaus widersprüchliche Situation, die erstmals 1890 von R. Altman richtig gedeutet wurde und die sich seither als eine der am besten belegten Theorien der Biologie etabliert hat:
Die Endosymbiontentheorie.
Im Laufe der Entwicklungsgeschichte scheint es mehrfach passiert zu sein dass eine Beute, die von einem Fressfeind sich einverleibt wurde aus welchem Grund auch immer nicht verdaut wurde sondert weiterlebte - mit vorteilhaften Folgen!
Zum ersten Mal passierte das wohl vor über einer Milliarde Jahren, als eine eukariotische Zelle sich ein Bakterium einverleibte, das vermutlich aus der Gruppe der sauerstoffatmenden Proteobakterien (Purpurbakterien?) stammte.
Im Laufe von Hunderten von Millionen Jahren optimierte die Evolution das gemeinsame Gespann so sehr, dass heute der Zellkern der eukariotischen Zelle sogar die DNS-Baupläne für lebensnotwendige Proteine dieser Mitochondrien enthält und ausliest..
Beide sind über die Produktion von ATP zu einem auf engste verzahntem Paar geworden, das getrennt nicht überlebensfähig wäre.
Bei den Plastiden ist die sache etwas komplizierter, hier müssen wir etwas weiter ausholen:
Vermutlich war es ein einzelnes Ereignis, als vor über einer Milliarde Jahre ein räuberischer farbloser eukariotischer Einzeller (der vielleicht schon Mitochondrien beinhaltete) sich eine Cyanobakterie einverleibte.
Aus ihrgendeinem Grund gelang es der Bakterie, sich der Verdauung zu widersetzen - befreien konnte sie sich allerdings nicht.
So blieb sie in ihrem Beutemacher; da dieser farblos und durchsichtig war wie die meisten Einzeller konnte die Cyanobakterie weiterhin Photosynthese betreiben und bei guter Lage sogar ihrem Träger einiges von dem dadurch gewonnenen Nährstoffen abgeben.
Im Gegenzug konnte sie ihrer Lage auch etwas positives abgewinnen:
Solange sie im Inneren ihres Wirtes lebte konnte sie von keinem anderen Räuber gefressen und dann doch noch verdaut werden.
Auch für den eukariotischen Beutemacher war die Situation nützlich:
Statt die Kuh zu schlachten und einmal Fleisch zu haben konnte er sie immer wieder melken.
Im Tausch gegen Schutz und Unterschlupf und vielleicht einige Mineralien, die er selbst nicht brauchte erhielt er energiereiche Nährstoffe, für die er selbst keinen Aufwand treiben musste, außer sich an helle Umgebung zu halten.
Ob es sich dabei um ein einzelnes Ereignis handelte oder ein mehrmaliges Ereignis zwischen zwei bestimmten Arten ist unklar.
Jedenfalls gehen folgende drei Gruppen auf diese Symbiose zurück:
-
Glaucocystophyceen: Unscheinbare Einzeller, ihre Plastiden (Cyanellen) enthält noch als einziger eine Bakterienzellwand aus Peptidoglucan.
-
Rotalgen: Hier besitzen die Plastiden (Rhodoplasten) noch den quasi originalen Antennenapparat der Cyanobakterien zum Lichtfang.
-
Grünalgen und höhere Pflanzen (Chloroplasten): Der lichtfangende Apparat (Antenne) ist stark weiterentwickelt.
Die Zurückführung auf dieses Ereignis war einer der Gründe, diese drei Gruppen zusammenzufassen und sie als eine der sechs Supergruppen in das neueste System aller Lebewesen nach Adl et alii einzuführen.
Sie firmieren dort als als Archaeplastida.
Das Leben und sein Stammbaum wäre heute viel einfacher und überschaubarer, wäre es bei diesem Ereignis geblieben.
Aber der Schutzraum, der eukariotische Unterschlupf und Wirt, wurde schließlich selbst zum Opfer eines anderen räuberischen eukariotischen Einzellers und das Cyanobakterium in ihm ebenso.
Wiederum gelang es nicht, das Beutetier zu verdauen und der Plastid, zu dem die Cyanobakterie mittlerweile geworden war überlebte mitsamt seinen ehemaligem Wirt.
Im Laufe vieler Generationen bildeten sich nun praktisch alle Zellbauteile des ersten Wirtes zurück - immerhin degeneriert DNA langsam aber stetig durch zufällige Kopierfehler falls kein Evolutionsdruck besteht.
Schließlich blieb von ehemaligen ersten Wirt nur die Zellhülle übrig, die sich schlicht als weitere Hülle über den Plastiden legt.
Bei Cryptophyceen konnte man zwischen den beiden Zellhüllen den Rest eines degenerierten Zellkerns nachweisen, vermutlich stammt er von einer Rotalge.

Die Gruppe der Lebewesen, die ein zweites mal einen Photosynthese-Symbionten erlangt haben (sekundäre Endocytobiose) sind in dem neuesten System aller Organismen von Adl et alii als Chromalverolata bezeichnet.
Dinoflagellaten sind oft zur Photosynthese fähig, müssen diese Ernährung durch Fressverhalten ergänzen, da sie nicht alle lebensnotwendigen Stoffe selbst bilden können.
Daher ist bei manchen Arten der originale Chloroplast zurückgebildet und funktionslos; diese Arten sind dann vollständig auf Beute angewiesen.
Durch das Fressverhalten haben sich manche Arten von Dinoflagellaten einen neuen Untermieter nutzbar gemacht;
das können ursprünglich Kalkalgen, Braunalgen oder Grünalgen o.a. gewesen sein.
Handelt es sich bei dem Clorophlasten vormalig um das Ergebnis einer sekundären Endocytobiose, so liegt nun eine dritte, tertäre Endocycobiose vor.
Da Dinoflagellaten ursprünglich bereits über einen Chloroplasten verfügen,
der noch in vielen Arten vorhanden ist und sich dieser als Rotalgenabkömmling identifizieren lässt, stehen die Dinoflagellaten nach wie vor in der Gruppe der Chromalveolata.
Plastiden und Mitochondrien sind genetisch so sehr verschieden, daß eine gemeinsamer Vorfahre derzeit ausgeschlossen wird.

|
Ophrydium v.:
Endosymbiose
|
Doch auch in der Gegenwart ist Symbiosen zwischen Algen und Tieren zu beobachten;
dabei sind die Algen (noch) genetisch autark und tauschen lediglich Stoffe mit dem Partner aus.
Der tierische Partner bezieht dabei ein Drittel bis knapp der Hälfte der Photosyntheseprodukte der Algen, die Alge erhält neben Schutz und Transport auch mineralischen Dünger (Stickstoff), der als Stoffwechsel-Endprodukt beim tierischen Partner anfällt.

|
sessiler Einzeller.
sym. Grünalgen
|
Auch sind Vielzeller bekannt, die zumindest eine Zeitlang auf die Verdauung und Verwertung der Algen verzichten, zumindest solange kein akuter Nahrungsmangel herrscht.
Während der ausgesetzten Verdauung nutzen sie die Assimilationsprodukte der Algen.
Die Meeresschnecke Elysia chlorotica nimmt die Plastiden ihrer Hauptnahrung, der Alge Vaucheria, in die eigenen Zellen auf und überlebt ohne Nahrung bei genügend Licht praktisch ohne eine Verkürzung der Lebenszeit.
Als Zoochlorellen bezeichnet man allgemein Algen, die im Inneren von Tieren leben, seien es nun Einzeller oder Vielzeller und mit ihnen eine dauerhafte, zum beiderseitigen Vorteil ausgerichtete Gemeinschaft eingehen.
Im Bild links: Ophrydium versatile, das Grüne Gallertkugeltierchen. Ein koloniebildendes Wimpertierchen, das in der Oberfläche einer gemeinsam gebildeten Gallertkugel lebt und sich von Algen und Bakterien ernährt.
Zahlreiche eingelagerte symbiontische Zoochlorellen färben der Körper grün.
Aber auch bei der Gattung Hydra findet sich eine Art, die Zoochlorellen einlagert.
Auf Safari mit der Lupe
Was nun folgen soll ist ein kleiner Überblick über die großen, oft augenfälligen Gruppen von Mikroorganismen.
Auch auf einer Safari findet man die Attraktionen nicht systematisch wie im Lehrbuch angeordnet sondern auf Nashorn folgt eine Horde von Geiern, dann ein Büffel und immer wieder mal ein Termitenbau.
Nun, unter dem Mikroskop ist das nicht anders.
Amöben

|
|
Amöbe.
|
Wer den Film "Der Blob" kennt wird das Schleim-Monster unter dem Mikroskop wiedererkennen:
Ein gallertiger Klumpen, der quasi durchsichtig ist und dessen Körper scheinbar keine feste Form kennt.
Zur Fortbewegung stülpt die Amöbe ein Scheinfüßchen aus, in das immer mehr Körpermaterial nachfließt, so dass der Körperschwerpunkt sich verlagert.
Schließlich geht das Scheinfüßchen (Pseudopodium) in dem nachfließenden Plasma wieder auf.
Zum Nahrungserwerb umfließt die Amöbe das Beutetier und bildet einen Hohlraum, der die Beute gefangenhält.
Der Hohlraum verengt sich und spaltet einen kleineren Hohlraum ab, der das miteingeschlossene Wasser nach außen abgibt.
Im Beutehohlraum lösen Verdauungsenzyme die Beute auf, die gewonnenen Nährstoffe gehen als Vakuolen in den Amöbenkörper über.
Im Mikroskop lassen sich der granulierte, körnige innere Bereich (Entoplasma) von einen schmalen äußeren Bereich unterscheiden, der glatt und glasig erscheint.
Von diesem äußeren Bereich werden die Scheinfüßchen gebildet.
Die genauen Abläufe sind noch nicht ganz geklärt, fest steht jedoch dass der Hauptakteur der Plasmabewegung ein im ganzen Körper verteiltes Netzwerk von Actin- und Myosinfilamenten ist,
also Molekülen, die auch in den Muskeln von Vielzellern die Bewegung erzeugen.
Gleichzeitig falten sich am Hinterende der Amöbe Kettenmoleküle zu engen Verbänden und verringern so das Volumen, während an der Spitze des sich bildenden Scheinfüßchens sich diese Kettenmoleküle entfalten:
Ein Druck auf das Zellplasma entsteht, der von Hinterende ausgehend das Plasma nach vorne treibt.
Da Plasma strömt dabei durch das Netzwerk der Actin- und Myosinfilamente, das zwar für das feine Endoplasma selbst durchlässig ist, nicht aber für die größerkörnigen Bestandteile des Endoplasmas und die Zellorganellen.
Da das Netzwerk an der Grenze zwischen Endo- und Ektoplasma besonders dicht ist, dürfte es die Ursache dafür sein dass Zellorganellen wie auch Nahrungspartikel nie das Endoplasma verlassen.
Ebenso bewirkt es dass das Ektoplasma klar ("gefiltert") erscheint und nicht körnig wie das innere Endoplasma.
Die Zellflüssigkeit fließt also durch den gesamten Zellraum, wobei sich das Ektoplasma am Hinterende von einer gelartigen Konsistenz zu einer weniger zähen Flüssigkeit (Kolloid) umwandelt, sich durch das Actin-Myosin-Filament nach voren schieben lässt.
Das im neuen Scheinfüßchen angekommene Endoplasma verwandelt sich im Randbereich der Amöbe wieder zurück in gelartiges Ektoplasma.

|
|
Amöbe.
|
Die grobe Fließrichtung ist dabei festgelegt, die Amöbe hat durchaus ein vorne und hinten - das Plasma sammelt sich stets an der Hinterseite (Uroid) und fließt durch die Körpermitte nach vorne.
Auch wenn es so aussieht als hätte die Amöbe keine feste Zellwand oder Hülle:
Im Elektronenmikroskop wird eine sehr dünne Membran sichtbar, die den Amöbenkörper von der Umwelt abgrenzt.
An ihrer Außenseite ist sie mit einer schleimartigen Härchenschicht überzogen (Mucoidschicht), die bei der Haftung am Substrat und beim Beutefang eine Rolle spielt.
Als lebender Teil der formveränderlichen Zelle kann sie jederzeit eingeschmolzen und neu gebildet werden.

|
|
Amöbe.
|
Im Amöbenkörper sind etliche Zellorganellen sichtbar.
Auffällig durch ihre Bewegung ist die kontraktile Vakuole, durch die überschüssige Flüssigkeit aus dem Zellinneren in die Umgebung geschafft wird.
Etwa alle 5 Minuten ist diese Pumpbewegung sichtbar.
Weiterhin finden sich ein scheibenförmiger Zellkern, Mitochondrien, zahlreiche kleine Kristalle in kleinen Vakuolen (das sind nicht versehentlich gefressene Staubkörner), Speichergranulae aus Fetten und Eiweißen und Nahrungsvakuolen.
Amöben ernähren sich räuberisch vor allem von Wimpertierchen und Geißeltierchen, aber auch von Kieselalgen und anderen lebenden Tieren.
(Sie sind der klassische Fressfeind der Pantoffeltierchen.)
Totes Material wird verschmäht.
Amöben können bis zu drei Wochen hungern, verlieren aber dabei einen Großteil ihrer Körpermasse (bis 90 %).
Die Vermehrung erfolgt ausschließlich durch Teilung, bei Zimmertemperatur alle 1 bis 2 Tage, bei ungünstigen Lebensbedingungen können Zysten als Dauerstadien gebildet werden.
Amöben sind relativ große vertreter der Protisten, Arten wie das häufige Amoeba proteus erreichen einen Durchmesser von etwa 600 Mikrometern (0,60 mm) , sind also bei genauerem Hinsehen mit blosem Auge gerade noch erkennbar.
Schalenamöben (Thecamoeben, Testaceen)

|
|
Schalenamöbe.
|
Schalenamöben sind die Schwestern der Nacktamöben;
sie leben in einer Schale, die aus nur einer Kammer besteht.
Wie sie bewegen sie sich kriechend mit Hilfe von Scheinfüßchen, also Ausstülpungen des Zellplasmas fort.

|
|
Schalenamöbe.
|
Chitinverwandte Substanzen verleihen der Schale Festigkeit, zum Teil wird auch Kieselsäure eingelagert.
Oft ist sie flach und rund, daher werden viele Schalenamöbenarten auch als Uhrglastiere bezeichnet.
Aber auch runde, gedrungene oder langgestreckte, keulenartige Formen kommen vor.
Von dort aus schieben sich ihre Scheinfüßchen nach außen, wobei sich je nach Art teils lappenartige Gebilde zeigen, teils aber auch eher dünne lange fadenförmige Strukturen, die nur aus Ektoplasma bestehen.
Die Zellorganellen wie auch Nahrungsvakuolen bleiben stets in der Schale.

|
|
Uhrglastierchen.
|
Wie Nackamöben ernähren sie sich räuberisch von kleinen Algen, Blaualgen, Bakterien und anderen kleineren Organismen.

|
|
Schalenamöbe.
|
Die Vermehrung dieser Einzeller erfolgt teils über eine simple Querteilung, bei der das Gehäuse auf die beiden Tochterzellen verteilt wird.
Bei manchen Arten tritt ein Teil des lebenden Zellmaterials aus der Schale heraus und beginnt vor der Teilung des gesamten Zellplasmas damit, eine neue Hülle aufzubauen.
Die Schalen von gestorbenen Thecamöben sind ziemlich dauerhaft und zersetzen sich nur langsam.
Sie sind in Wasserproben mitunter recht häufig anzutreffen und zumindest bei den Uhrglastierchen leicht zu erkennen.
Rädertierchen

|
|
Rädertierchen
|
Rädertierchen (Rotatorien, Rotifera) sind kleine vielzellige Tierchen, deren charakteristisches gemeinsames Merkmal ein Räderorgan an der Stirnseite darstellt, das sie zur Fortbewegung und zum Nahrungserwerb nutzen.
Das geringe Auflösungsvermögen der ersten Mikroskope, durch die diese Tiere beobachtet wurden führte zu dem Eindruck, dieses Kopforgan stelle eine Art Propeller das oder jedenfalls etwas schnell rotierendes.

|
|
Rädertierchen
|
Tatsächlich aber besteht das Räderorgan aus einem festsitzenden Kreis von Wimpern, die ein darunter umlaufender Nervenimpuls jeweils aktiviert.
Die Wimpern schlagen dann in schneller Abfolge peitschenförmig aus und bewegen so das umliegende Wasser.
Dabei wird Nahrung herangestrudelt, in die Mundöffnung eingestrudelt oder das Rädertierchen nutzt den entstehenden Rückstoß zur Fortbewegung.
Rädertierchen sind verschiedengestaltig gebaut, besonders das Räderorgan ist je nach Art charakteristisch und mitunter eindrucksvoll.
Aber stets folgen sie dem gleichen Bauplan, der den Körper in eine Kopfpartie, einen Rumpf und eine Fuß einteilt.

|
|
Herz-Rädertier.
|
Der Kopf ist Träger des Räderorgans, je nach Art kann er mitunter fast vollständig eingezogen werden.
Die vom Räderorgan eingestrudelte Nahrung wird durch die Speiseröhre in den Kaumagen befördert, wo mehrere harte Strukturen gegeneinander reiben um die Nahrung zu zerkleinern.
Bau und Härte dieser Gegenstücke ist typisch für die Art der bevorzugt aufgenommenen Nahrung und ermöglichen bei den unterschiedlichen Arten eine Vielzahl von Ernährungsweisen.
Im daran folgenden Magen wird die Nahrung verdaut, der zwischen Rumpf und Fuß befindliche After scheidet die Reste aus.
Bei manchen Arten, die praktisch ausschließlich im freien Wasser leben ist der Fuß rückgebildet.
Diese Rotatorien erscheinen eher sackförmig.

|
|
Herz-R.: Kolonie.
|
Rädertierchen besitzen ein lichtsensorisches Organ, das paarig vorhanden ist und sich zwischen Räderorgan und Kaumagen befindet. Oft ist es durch seine orange Färbung gut zu erkennen.

|
|
Bdelloides R.
|
Ein komplexes System von Längs- und Quermuskeln ermöglichen eine Fülle von Bewegungen und erlaubt es den Rotatorien sich geschmeidig sowohl durch das Wasser auch als durch dichtes Algenfadengestrüpp zu bewegen.
Die weitaus meisten Arten von Rotatorien leben von planktischen Algen, Bakterien und Detritus.
Nur wenige Arten sind zu einer räuberischen Lebensweise übergegangen (zB. Asplanchna).
Der Fuß dient als Schwimmruder beim freien Schwimmen, als Haftorgan mittels Klebezellen und zur Fortbewegung von Arten, die sich spannerartig fortbewegen.
(Spannerartig: Der Fuß haftet am Substrat, Streckbewegungen schieben den Vorderkörber nach vorne; dann löst sich der Fuß vom Substrat und wird nachgezogen.)
Manche Arten bevorzugen eine weitgehend sesshafte Lebensweise, dann ist der langgestreckte Fuß oft mit einer durchscheindenden Gallertschicht umhüllt, in die sich das Rädertier zurückziehen kann.

|
|
Gallerthülle.
|
Oft sitzen Rotatorien wenige Sekunden an einer Stelle, strecken und kontrahieren sich dabei unermüdlich und strudeln emsig nach Nahrungspartikeln.
Dann lösen sie sich und schwirren umher zu einem anderen Standort.

|
|
Mützenrädertier
|
Die Vermehrung bei Rädertieren erfolgt fast immer durch Parthenogenese, d.h. unbefruchtete Weibchen bringen Töchter hervor, die über das gleiche Erbgut verfügen wie ihre Mütter (Klone).
Nur gelegentlich, von bestimmen Umweltfaktoren ausgelöst, bilden sich Männchen und befruchtungsfähige Weibchen.
Sie zeugen Dauereier, die Mangelperioden (Winter, Nahrungsknappheit, Trockenheit) überstehen.
Bei vielen Arten sind auch adulte Tiere dazu in der Lage, in ein Dauerstadium überzugehen, indem sie einen Teil der Körperflüssigkeit abgeben und sich mit einer dicken Schutzhülle umgeben.
In Mitteleuropa sind etwa 500 Arten von Rädertierchen bekannt, die in die Bdelloida (langgestreckt, spannerartige Fortbewegung) und die Monogononta (sackartig, meist freischwimmend) eingeteilt werden.
Wimpertierchen

|
|
Wimpertierchen.
|
Von allen Protisten stellen die Wimpertierchen die am höchsten entwickelte Gruppe dar.
Diese Einzeller bieten eine Überfülle an Arten mit unterschiedlichem Körperbau und Lebensweise, ebenso stellen sie zahlenmäßig eine ungewöhnlich häufige Gruppe dar,
so dass man sie wohl als die erfolgreichsten Vertreter der Protisten bezeichnen darf.
Ihnen gemeinsam ist ein Wimpernkleid, das den Körper ganz oder zumindest teilweise bedeckt und mit dessen Hilfe Nahrung herbeigestrudelt und die Fortbewegung bewerkstelligt wird.
Bei weiter entwickelten Arten ist das Wimperkleid reduziert oder zum Teil modifiziert.
Bei den Lauftierchen sind Wimpern zu kräftigen Zirren verschmolzen, bei Glockentierchen und anderen finden sich Wimperbänder die in festgelegten Mustern den Körper umlaufen.
Die Wimpern (Cilien) erzeugen einen Rückstoß indem sie nach der Ruderbewegung bei der Rück-Bewegung abknicken.
Sobald sie die vordere Ausgangslage erreicht haben richten sie sich wieder auf und sind zum nächsten Ruderschlag bereit.
Die Wimperspitze beschreibt dabei eine Kreisbahn, deren Flächeninhalt ein Maß für den Vortrieb ist.
Die Wimpern stehen in Längsreihen, die wiederum in einzelne kleine Wimperfelder unterteilt sind.
Da die Cilien nicht allesamt synchron schlagen sondern leicht phasenversetzt, sieht man bisweilen ein Wellenmuster den Körper entlanglaufen, das durch diejenigen Wimpern gebildet wird, die gerade die gleiche Bewegung vollführen.
Mit der Anordnung der Cilien haben diese Muster nichts zu tun, sie sind weitgehend variabel und resultieren aus der Schwimmrichtung und dem momentanen Schwimm-Manöver des Wimpertierchens.

|
|
Stentor
|
Mit dem hochflexiblen Antriebskonzept der Bewimperung und der Koordination der vielen einzelnen Wimpern schaffen es die Ciliaten, vorwärts, rückwärts, in Kurven und mit unterschiedlichen, angepassten Geschwindigkeiten sich fortzubewegen,.
Die Wimpern entspringen je einem Basalkörper, der dicht unter der Zellmembran liegt. Ihr innerer Aufbau ist im ganzen Tierreich fast universell:
Zwei getrennte Mikrotubuli (röhrenförmige Mikrostrukturen aus Protein) in der Zentralachse der Wimper sind umgeben von neun kreisförmig angeordneten, je doppelt angelegten Mikrotubuli;
Längsverschiebungen der Tubuli gegeneinander führen zu einer einseitgen Längenveränderung und damit der Krümmung der Cilie.
Viele Wimpertierchen verfügen über einen Abwehrmechanismus, der sie vor Fressfeinden wie der Amöbe schützt.
Diese Trichozysten entstehen im Zellplasma als kleine Bläschen (Vesikeln) und werden unter der Oberfläche der Wimpertiere verankert, wo sie auf ihren Einsatz waarten.
Sie bestehen aus einem länglichen Geschossteil, der von einer Schutzhülle umgeben ist, und einem Auslösemechanismus.
Bei Gefahr schleudern die Trichocysten einen Faden nach außen, der in der Vielzahl einen regelrechten Pelz ergibt, unter dem das Wimpertierchen schwer faßbar wird.
Dieser Vorgang ist erstaunlich schnell und dauert nur Millisekunden; diese Geschwindigkeit kann vermutlich nur durch einen Quellungsvorgang erreicht werden, bei dem sich das Geschossteil auf das zehnfache seiner ursprünglichen Länge ausdehnt.
Bei manchen Arten sind die Fäden vermutlich toxisch (Trichiten, Toxicysten), teils werden statt Fäden auch Schleime ausgestoßen.

|
|
Wimpertierchen.
|
Zur ungeschlechtlichen Vermehrung teilen sich Wimpertierchen quer (Ausnahme: Glockentiere, dort Längsteilung).
Dabei entsteht aus dem vorderen und dem hinteren Ende des Tieres je ein neues Individuum.
Die vordere Tochterzelle (Proter) übernimmt dabei den unsprünglich vorhandenen Mundapparat des Muttertieres, das hintere neue Tier (Opisthe) bildet einen neuen Mund aus.
Dabei sind die beiden Tochtertiere identisch und nicht spiegelbildlich wie sich das bei der Längsteilung der Flagellaten ergibt.

|
|
Wimpertierchen.
|
Auch der Austausch von Genmaterial ist möglich, wenn auch unter hohem Risiko.
Bei diesem Vorgang (Konjugation) erfolgt allerdings keine Vermehrung.
Dazu lagern sich die beiden Partner eng aneinander und verschmelzen an der Berührungsstelle.
Wimpertierchen besitzen zwei Zellkerne, von denen der weit größere (Makronucleus) nur die Stoffwechselvorgänge steuert.
Er wird aufgelöst.
Nun wird kleinere Kern (Mikronucleus) aktiv, der bei Sexualvorgängen die entscheidende Rolle spielt.
Zunächst teilt er sich zwei Mal, so daß vier Tochterkerne entstehen, die je über einen halbierten Chromosomensatz verfügen.
Drei dieser Kerne zerfallen wieder, der übrige Kern teilt sich ein weiteres Mal.
Von den nun paarig vorhandenen Kernen der beiden Partner wird je einer zum anderen Partner entsendet, sie wandern über eine Plasmabrücke in das Innere des Partners.
Ist der Tausch vollzogen, vereinen sich Wanderkern und stationärer Kern zum vollständigen Chromosomensatz und die Partner trennen sich wieder.
(Das ist in etwa der grobe Vorgang, der je nach Art abgewandelt und variiert ist.)
Etwa die Hälfte der Individuen stirbt direkt nach der Konjugation - ein hoher Preis, doch scheint Nutzen des Genaustausches so groß zu sein dass es die Sache wert ist.
Wimpertierchen werden in die zwei Gruppen Postciliodesmatophora und Intramacronucleata unterteilt, wobei die erstere eine deutlich geringere Artenzahl umfaßt.
Das Charakteristikum der Intramacronucleata ist ein Großkern (Macronucleus) der von Microtubuli, röhrenförmige Proteinstrukturen, unterteilt ist.
-
Postciliodesmatophora
-
Stentor

|
Stentor mit
symbiont. Algen
|
Das Trompetentierchen Stentor ist ein Prachtstück von einem Einzeller.
Mit 2 mm Länge, festsitzend ausgestreckt noch deutlich mehr erreicht es eine enorme Größe.
Als Süßwasserbewohner besiedeln sie schattige Uferzonen von langsam fließenden oder stehenden Gewässern mit reichem Pflanzenbewuchs.
Meist sitzen sie auf einem planzlichen Untergrund und ernähren sich von Algen, Bakterien aber auch Kieselalgen, Schwebealgen und anderer Beute, die sie in ihre Mundöffnung einstrudeln.
In Mitteleuropa sind etwa eine Hand voll Arten bekannt, darunter auch das Grüne Trompetentier, das durch eingelagerte symbiontische Algen grün gefärbt ist, oder das Blaue Trompetentier.
-
Blepharisma sp.
")
|
|
Blepharisma (?)
|
Blepharisma, speziel Blepharisma japonicum stellt einen in Forschungs- und Schulkreisen beliebten Einzeller dar, da er mit über einem Millimeter Länge eine imposante Länge erreicht und mit blosem Auge sichtbar ist.
Dazu ist er durch das Pigment Blepharismin charakteristisch rosa oder rötlich gefärbt.
Außerdem sind sie ähnlich wie das bekannte Pantoffeltierchen leicht zu halten und zu züchten; Kulturen sind bei Ausrüstern für den Biologieunterricht beziehbar.
In flachen Gefäßen (Petrischalen) lassen sie sich bei 24°C an einem dunklen Standort halten.
Ein eingebrachter Kohlrübenwürfel dient als Nahrungsgrundlage der Futterbakterien, allerdings begnügen sich gerade die größeren Arten von Blepharisma nicht ausschließlich mit Einzellern sondern benötigen auch kleinere Eukarioten als Futtertiere.
Bei Nahrungsmangel entsteht Kannibalismus, der bei den Überlebenden Riesenwuchs hervorbringt.
Auch die Alleinfütterung mit bestimmten Wimpertierchen führt zu Gigantismus, der Individuen von einem Millimeter Größe erzeugt.
-
Intramacronucleata
- Holotricha

|
|
Paramecium.
|
Hier finden sich längliche oder runde Formen, deren Wimpernkleid gleichmäßig über den Körper verteilt sind.
An der (in Fahrtrichtung) rechten Seite der Mundöffnung ist eine Reihe von Wimpern zu einer Membran, einem Bändchen, verschmolzen welches in wellenförmigen Bewegungen die Nahrungsaufnahme unterstützt (Mundsegel).
Vor einiger Zeit galt diese Gruppe noch als Verwandschaftsgruppe; heute gilt dieser Begriff nur noch als Bezeichnung für die Erscheinungsform, also eben vollständig bewimpert.
Das wohl bekannteste holotriche Wimpertierchen ist das Pantoffeltierchen.
Mehr Information zu Haltung und Zucht gibt es hier.
-
Peritricha: Glockentiere

|
|
Glockentierchen.
|
Bei diesen Wimpertierchen ist das Vorderende des Körpers zu einer runden Scheibe (Peristom) abgeplattet.
Am Scheibenrand entlang läuft spiralig linksgewunden eine Wimperreihe, die den Zellmund teils mehrfach umrundet.
Das innere Ende der Wimperreihe mündet in eine seitlich gelegene Öffnung in der Scheibe, den Mundtrichter (Zellmund, Cytostom).
Dort sammelt sich die Nahrung. Sobald die Menge eine Grenzgröße erreicht hat, wird sie als Vakuole zur Verdauung in das Körperinnere aufgenommen.
Glockentierchen ernähren sich in der Hauptsache von Bakterien.
Der Wasserstrom, den die Wimpern erzeugen lässt sich gut beobachten sobald sich einige Partikel in ihm mitbewegen.
Der restliche Körper ist meist nicht bewimpert.
Die meisten Glockentiere sind festsitzend und heften sich mit einem Stiel an das Substrat, was ihnen mitunter das Aussehen einer Tulpe verleiht.
Dieser Stiel wickelt sich bei unerwünschten Reizen (Berührung, Erschütterung) ruckartig zu einer Spirale ein und zieht das Glockentier schnell aus der Gefahrenzone.
Gleichzeitig wird das Mundfeld eingezogen.
Wird ein Glockentierchen von seinem Haftpunkt abgerissen oder ist ein neuer Standort erstrebenswert, so kann es sich mit Hilfe der Wimpern auch im freien Wasser zügig fortbewegen.

|
|
Kolonie.
|
Glockentiere sind aber auch in der Lage, relativ schnell Schwärmerstadien zu bilden.
Dabei werden die Wimpern am Vorderende abgebaut und es entstehen neue Fortbewegungswimpern am hinteren Ende (Schwimmringe).
Sobald sich das Tier vom Stil gelöst hat, schwimmt es rotierend und mit den Hinterende voran.
Die ungeschlechtliche Vermehrung erfolgt über Längsteilung.
Dabei beginnt die Teilung am Mundfeld und läuft nach unten; wird auch der Stiel geteilt und der gemeinsame Aufsitzpunkt erhalten ist der erste Schritt zu einer Koloniebildung getan.
Bei manchen Arten teilen sich mehrere Tiere einen verzweigten Stiel.
Die Tochterzelle kann sich aber auch als mobiler Einzeller mit Schwimmring ausbilden.
Die meisten Glockentierarten sind festsitzende Bakterienfresser und bilden die Ordnung Sessilida.
Einige wenigen mobilen Arten (Ordnung Mobilida) verfügen als freischwimmende Ektoparasiten über ein spezielles Anhaftungsorgan.
-
Spirotricha : Membranellentiere
")
|
|
Euplotes sp. (?)
|
Hier führt das oft stark ausgeprägte Mundsegel in einer rechtsdrehenden Windung zum Mund.
Mitunter führt das Band über den halben Körper, beginnend an einer Seite über das Vorderende hinweg bis zur Mundöffnung im hinteren Teil der anderen Seite.
Einige Arten (Hypotrichia) verfügen über stabile, kräftige Zirren, die aus verschmolzenen Wimpern entstanden sind und sich auf der Bauchseite befinden.
Mit ihrer Hilfe scheinen sie über das Substrat zu laufen (Lauftierchen).
Auch ermöglichen sie gezielte Tast- und Suchgewegungen.
Eine bekanntere Gattung hier: Euplotes.
-
Sauginfusorien
Bei diesen Wimpertierchen sind Mundöffnung und Wimperkleid vollständig zurückgebildet.
Sie sitzen auf einem kurzen, nicht kontraktilem Stil auf, meist als Aufsitzer auf anderen, festsitzenden Organismen.
Sauginfusorien (Suctoria) erbeuten ihre Nahrung nicht strudeld, sondern haben sie sich auf eine trickreiche Variante verlegt.
Statt einem Wimperkleid verfügen Sauginfusorien über etliche Saugtentakeln, die aus dem Körper ragen und die an der Spitze mit einem kleinen Knöpfchen versehen sind.
Sobald eine potentielle Beute einen Knopf berührt bleibt sie daran hängen, da sich der Knopf in der "Haut" (Pellicula) des Opfers verankert.
Fluchtversuche führen nur noch zu weiteren Berührungen mit anderen Knöpfchen.
Der Suctor injiziert nun lähmende Substanzen und Verdauungsenzyme über die Fangtentakel in das Opfer;
schließlich saugt er es durch die Tentakel vollständig aus, so dass von seiner Beute nur mehr eine leere Hülle übrig bleibt.
Sonnentierchen

|
|
Sonnentierchen.
|
Sonnentiere (Heliozoa) sind einzellige Organismen mit einem kugelförmigen Körper, von dem namensgebend viele dünne, gerade Scheinfüßchen radial allseitig abstehen, eben wie Sonnenstrahlen.
Der Körper ist durchsichtig wie bei Amöben und lässt bisweilen die letzte Mahlzeit des Tieres erkennen.
Die Scheinfüßchen (Axopodien) dienen nicht der Fortbewegung, sondern dem Beutefang.
Es besteht aus Zellplasma, das entlang eines stabilisierenden, mittig gelegenen Zentralstäbchens entlang ständig auf und ab läuft.
Bei vielen Arten ist der Zellkörper ungeschützt, bei anderen von einer Schleimhülle oder auch von selbsterzeugten oder angelagerten Außenskeletten aus Chitin, Kieselsäure oder Kieselalgenschalen umgeben.
Manche beherbergen einzellige Grünalgen und profitieren von deren Stoffwechselprodukten.
Die meisten Arten von Heliozoa schweben frei im Wasser und betreiben keine aktive Fortbewegung (womöglich dienen die Axopodien einem längeren Schweben).
Andere Arten sitzen auf kurzen Stielen auf dem Substrat auf.
Aktive Fortbewegung ist nur als langsamen Rollen mit hilfe der Axopodien bekannt.
Sonnentiere ernähren sich räuberisch von Kieselalgen, Schwebealgen, Wimpertierchen geeigneter Größe und auch kleinen Rädertierchen.
Dabei erbeuten sie Organismen, die die Spitzen der Axopodien berühren und dort wie gelähmt kleben bleiben.
Dabei bewirken Extrusomen, die als kleine Körnchen stets in Plasmastrom des Axopoden mitschwimmen, einen Klebeeffekt, der von mechanischen oder biochemischen Reizen, die von einem Beutetier ausgehen, ausgelöst wird.

|
Sonnentierchen:
Nahrungsaufnahme.
|
Das Scheinfüßchen, an dem die Beute hängt wird nun langsam eingeschmolzen und das Zentralstäbchen abgebaut und verkürzt, während das frei werdende Plasma an der Axopodien-Spitze die Beute überwölbt und einhüllt.
Die so entstehende Nahrungsvakuole verschwindet schließlich im Körper des Sonnentieres, die Inhalt wird verdaut.
Unverdauliche Reste werden als Vakuolen an die Umgebung abgegeben, mitunter ist das als Bläschen an der Oberfläche erkennbar.
Vermutlich fangen die Extrusomen ständig kleine Beute wie Bakterien und verfrachten diese in den Körper; die Axopodien fungieren dabei - ohne sich zu verkürzen - eher wie ein Förderband.
Mitunter vereinen sich zwei oder mehr Tiere zu einer Freßgemeinschaft, indem sie ihre Körper zum Teil verschmelzen und so eine größere Beute zu überwältigen.
Nach der Verdauung, sobald die Beute in mehrere Nahrungsvakuolen umgewandelt und damit teilbar ist, trennen sie die Tiere wieder.
In freier Natur sind Sonnentiere weit verbreitet und treten ganzjährig in stehenden oder langsam fließenden Gewässern auf.
Häufig sind sie vor allem aber im Sommer im Schwimmdetritus, zwischen Wasserpflanzen und besonders in Algenwatten, die an der Wasseroberfläche treiben wo sie von Wärme und hohem Sauerstoffgehalt profitieren.
Sonnentiere überdauern Trockenheit problemlos in einer undurchlässigen Schutzhülle (Schutzzysten).
Sobald sie sich wieder in nasser Umgebung befinden läuft der Stoffwechsel wieder an.

|
|
Sonnentierchen.
|
Die Vermehrung erfolgt ungeschlechtlich über Teilung;
die geschlechtliche Vermehrung setzt nur bei Ressourcenknappheit oder anderen ungünstigen Lebensverhältnissen ein.
Dabei bildet sich innerhalb des Sonnentieres eine Mutterzyste, aus dem Tochterkugeln abgeschnürt werden.
Diese Tochterkugeln teilen sich zwei mal und verfügen dann über einen einfachen (haploiden) Chromosomensatz.
Diese verschmelzen paarweise und bilden so eine befruchtete Eizelle (Zygote) mit doppeltem, diploidem Chromosomensatz.
Diese Zygote bildet eine dicke Schutzschicht und verbringt eine Ruhephase, nach der sie auskeimt.
Sonnentierchen stellen keine Gruppe von Arten dar, die auf einen gemeinsamen Vorfahren zurückgeht.
Vielmehr ist sie - wie andere Gruppen von Protisten, die aufgrund gemeinsamer äußerlicher Merkmale zusammengruppiert wurden - eine Gruppe von nicht näher verwandten Tieren, die eine gemeinsame Organisationsstufe erreicht haben,
also unabhängig voneinander ein funktionierendes Konzept zum Überleben entwickelt haben und deswegen sehr ähnlich aussehen.
(Nach Adl et alii entstammen die Sonnentierchen teils den Rhizaria, teils den Chromalveolata.)
Algen - Grünalgen und andere Algen
Unter Algen stellt man sich üblicherweise die fädigen, watteartigen Fladen vor, die im Spätsommer in nährstoffhaltigen Weihern und Seen an der Oberfläche schwimmen.
Diese Fadenalgen stellen aber nur einen kleinen Teil der unübersichtlichen und formenreichen Gruppe der Grünalgen dar.

|
|
Grünalge.
|
Allgemein wird unter einer Alge ein Organismus verstanden, der dauerhaft Photosynthese betreibt und bei der Fortpflanzung über keine Embryonalstadium verfügt.
Wenn man bedenkt, dass sich Algen als eukatiotische Einzeller aus einer Verbindung von einem Einzeller, der sich als Räuber, also tierisch ernährt und einer Cyanobakterie hervorgegangen sind,
so wird klar dass Algen keineswegs die festsitzende, unbewegliche Lebensweise betreiben wie man das von Landpflanzen kennt.
Stattdessen verfügen sie oft, wie der Einzeller aus dem sie entstanden sind, über eine oder mehrere Geißeln, die der aktiven Fortbewegung dienen.

|
4-fach Kolonie
mit Gallerte
|
Je nach Organisationshöhe oder Lebensphase ist die Geißel zurückgebildet und die Zellen haften aneinander und bilden (scheinbar) ein Gewebe.
Die einfachste Organisationsstufe bilden dabei einzellige, frei lebende begeißelte Algen.
(Man kann sie auch zu den pflanzlichen Flagellaten zählen.)
Sie stehen dem ursprünglichen Einzeller wohl noch an nächsten.

|
Scenedesmus sp.
Gürtelalge.
|
In der nächsten Stufe schließen sich Algenzellen zu einer Kolonie von mehreren oder vielen begeißelten Zellen zusammen (monadoide Stufe).
Die Kolonie ist von einer Gallerte umgeben. (Die bekannte Gattung Volvox, das gerne als eines der sieben Weltwunder der Mikrobiologie bezeichnet wird, gehört hierzu.)

|
Pediastrum sp.
Zackenrädchen
|
Als weiteren Schritt (die Einteilung ist subjektiv) verlieren im weiteren Verlauf der Evolution die koloniebildenden Zellen ihre Geißel.
Es entsteht die capsale Stufe; ihr Kennzeichen sind Kolonien von nach wie vor eigenständigen, aber unbegeißelten Algenzellen.
(z.B. Scenedesmus, Pediastrum).
Viele Arten, die in der Wachstumsphase auf Geißeln verzichten, bilden diese an den Geschlechtszellen aus um räumlich weiter zu streuen.
Verzichtet eine Alge in ihrer evolutiven Entwicklung auf Koloniebildung und schützt sie sich stattdessen mit einer widerstandsfähigen Schicht, so stellt sie diese Form die kokkale Stufe dar.
Hierzu zählen etwa die Kieselalgen.
Fadenalgen bilden sich wenn eine Alge sich stets in eine gleichbleibende Richtung teilt und dabei eine gemeinsame Kontaktfläche zur Tochterzelle erhalten bleibt.
Die watteartigen Fladen, die an der Wasseroberfläche flottieren finden sich hier (trichale Stufe).
Verzweigungen sind natürlich auch möglich.
Charakteristisch sind die gut sichtbaren Querwände, die in regelmäßige Abständen den Fadenkörper strukturieren.
Solche Algen sind nun keine echten Einzeller mehr, sie bilden einen zusammenhängenden Körper, den Thallus.
Wachsen nun die Zellen bei der Zellteilung nicht nur entlang einer Linie sondern in alle drei Raumdimensionen, so entstehen gewebeartige Körper, die mitunter beachtliche Dimensionen aufweisen können.
Die Thalli von Rotalgen und Braunalgen werden mitunter viele Meter lang und bilden manchmal regelrechte Unterwasserwälder.
Sobald ein Pflanzenkörper in Spross, Blatt und Wurzel unterteilt werden kann, also echtes Gewebe aufweist, und sobald ein Embryonalzustand vorhanden ist zählt er nicht mehr zu den Algen sondern zu den höheren Pflanzen.
-
Rotalgen:
Meist marin, meist vielzellig. Ohne Geißeln. 1,4 bis 1,2 Milliarden Jahre alt.
-
Gelbgrüne Algen (Xanthophyceae):
Farblich gleichen sie den Grünalgen;
während aber Grünalgen über zwei gleich lange Geißeln verfügen sind die beiden Geißeln der Gelbgrünen Algen unterschiedlich lang.
Ebenso groß ist der Unterschied zu anderen Gruppen hinsichtlich ihrer Körperchemie: Gelbgrüne Algen nutzen Öle als Reservestoff.
Manche Arten besitzen keine Zellwand, zumindest zeitweilig leben sie wie eine winzige Amöbe.
Andere Arten schließen sich zu Verbänden zusammen, die zu einer einzigen überdimensionalen Zelle mit zahllosen Zellkernen verwächst (Botrydium sp.).
Auch fädenförmige Strukturen kommen vor.
In der Regel geschieht die Fortpflanzung auf vegetativem Wege, also durch Teilung.
Geschlechtliche Fortpflanzung ist nur bei Vaucheria bekannt.
Die Artenzahl beträgt etwa 400, häufige Arten gibt es wohl nicht.
-
Goldbraune Algen (Chrysophyceae):

|
Dinobryon sp.
Becherbäumchen.
|
Goldbraune Algen leben meist als begeißelte Einzeller, Koloniebildung ist selten.
Von den beiden Geißeln ist die eine nach vorne, die andere, die am gleichen Ursprungsort ansetzt, seitlich gerichtet.
Die nach vorne gerichtete ist mit kleinen seitlich abzweigenden Härchen ausgestattet (Flimmergeißel).
Um die verschiedenen Algen zu unterscheiden ist der eingelagerte Plastid und seine Herkunft nicht der einzige Königsweg.
Auch der Reservestoff, den die Algen bilden um sich über schlechte (lichtarme) Zeiten zu retten ist ein taugliches Kriterium, um die verschiedenen Arten einzuordnen und in dem vermeintlichen Gewirr ein System zu etablieren.
Währen die Grünalgen typischerweise Stärke als Reservestoff nutzen (andere Gruppen nutzen Öle) findet man in Goldbraunen Algen Chrysolaminarin, ebenfalls ein Kohlenhydrat, das aber anders aufgebaut ist als die Stärke der Grünalgen.
(Chrysolaminarin wird auch von Kieselalgen benutzt.)
Oft sind diese Einzeller in der Lage, ihre Geißeln abzuwerfen und sich zu einer Kolonie zusammenzufinden.
Da sie sich in der gemeinsamen Gallerthülle auch teilen können entstehen mitunter große Ansammlung von schleimigen Polstern.
Ein unverwechselbarer Vertreter der Goldbraune Algen ist Dinobryon, ein Gehäuse- und Koloniebildner.
Nach einer Teilung verbleiben die Tochterzellen nahe des Gehäuses der Mutterzelle.
-
Cryptophyta:
Kaum beachtet bilden sie trotzdem ein wichtiges Standbein im Ökosystem.
Durch ihre geringe Größe dienen sie vielen anderen Einzellern als Nahrung.
Cryptophyta haben ihren Plastiden sekundär erworben; er stammt also von einer eukariotischen Alge, die bereits einen Plastiden beherbergte.
Sie wurde von dem Vorläufer der Cryptophyta aufgenommen und nicht verdaut.
Im Laufe der Evolution bildete sich die Alge soweit zurück dass nurmehr ihre Zellmembran um den Plastiden herum übrig blieb - und ein kleiner, stark zurückgebildeter Zellkern, ein Nucleomorph.
Molekulargenetische Untersuchungen konnten hier ganz konkret nachweisen, dass es sich hier um den Rest eines Rotalgen-Zellkerns handelt - ein starker Beleg für die Endosymbiontentheorie.
In der Regel stellt diese Gruppe kleine Arten, die mit zwei unterschiedlich langen Geißeln ausgestattet sind und einzeln ohne Koloniebildung leben.
Wer also unter dem Mikroskop kleine, grünliche Organismen zu erkennen glaubt, die flink und unablässig heumwirbeln: Das könnten Cryptophyten sein.
-
Grünalgen:
Von allen im Süßwasser lebenden Algen stellen die Grünalgen mit gut der Hälfte die meisten Arten.
Weltweit gibt es etwa 7000 Arten (einschließlich der marinen Arten).
Grünalgen lassen sich in zwei große Gruppen gliedern: Charophyta und Chlorophyta.
Die Charophyta umfassen Zieralgen, Jochalgen, die Armleuchteralgen (die bereits so hoch organisiert ist dass sie auf den ersten Blick einer Pflanze ähnelt) und die evolutiv daraus hervorgegangenen Höheren Pflanzen.
Zieralgen und Jochalgen werden zusammenfassend auch als Schmuckalgen bezeichnet.
Schmuckalgen sind nun die artenreichste Gruppe und stellen etwa 4000 Arten, die weltweit im Süßwasser vorkommen.
-
Zieralgen

|
|
Cosmarium botrytis.
|
In dieser Gruppe finden sich viele optisch sehr ansprechende Arten.
Stets sind sie zweigeteilt, die beiden Hälften sind über einen mehr oder weniger breiten Kanal verbunden, der eine Plasmabrücke zwischen den beiden Zellhälften bildet.
In diesem Kanal befindet sich der Kern, in den beiden fast perfekt symmetrischen Hälften je ein Plastid.
Bei der Zellteilung halbiert sich die Algenzelle am Kanal, die beiden Tochterzellen müssen also je eine Hälfte nachbilden.
Ist eine Hälfte der Zelle deutlich kleiner als die andere so lässt sich daraus auf eine jüngst erfolgte Teilung schließen.

|
Closterium moniliferum
Mondsichel.
|
Zur geschlechtlichen Fortpflanzung legen sich zwei Algen unterschliedlichen Fortpflanzungstyps aneinander und bilden eine gemeinsame schützende Plasmaschicht aus.
Dann werden die sich berührenden Zellwände aufgelöst und die beiden Zellkerne wandern aufeinander zu und verschmelzen.
Diese Verschmelzung wird nicht sofort rückgängig gemacht, die beiden Zellen bleiben bis auf weiteres verschmolzen und bilden manchmal eine gemeinsame Schutzschicht aus.
Das Verschmelzungsprodukt durchläuft nun eine Ruhephase und beginnt nach deren Ende die neue Vegetationsperiode mit einer Reifeteilung, die den doppelten Chromosomensatz zum normalen, einfachen Chromosomensatz reduziert.
-
Jochalgen

|
|
Spirogyra.
|
Jochalgen verfügen über die gleiche Fähigkeit zur Konjungation wie ihre Schwestern, die Zieralgen.
Allerdings bilden sie fädige, unverzweigte Formen aus.
Kommt es zur sexuellen Reproduktion, so lagern sich zwei Fäden parallel aneinander und bilden eine Brücke aus.
Bilden mehrere Zellen zweier Fäden eine Brücke, so ähnelt das Ergebnis entfernt einer Leiter, bei der an langen Holmen gelegentlich eine Sprosse erscheint.
Die verbundenen Zellen können bei fädigen Strukturen nicht verschmolzen bleiben ohne den Zusammenhalt des Fadens zu gefährden;
Daher wandert in der Regel der Zellinhalt des einen Partners in die Zellhülle des anderen und verbleibt dort.
Er hinterläßt in seinem angestammten Faden eine leere Zellhülle.

|
|
Algen.
|
Interessant dabei: Es ist immer der gleiche Faden, dessen Einzeltiere zum Partner hinüberwandern bzw. in der angestammten Zellwand verbleiben.
Da die Individuen eines Faden genetische Klone sind und es wohl keinen Zufallsmechanismus gibt, der festlegt, welcher Partner wandert, liegt der Verdacht nahe, die Rolle könnte genetisch festgelegt sein, also es könnte eine Art männliche oder weibliche Algenzelle geben.
Tatsächlich aber verhält sich der gleiche Faden unterschiedlich, wenn er mit unterschiedlichen anderen Fäden eine Verbindung eingeht.
Algen sind also nicht männlich oder weiblich, ihre Rolle ist relativ zum Partner definiert.
Einige Arten verschmelzen bei geeigneter Lage mit Zellen des gleichen Fadens.
Bekannt ist die Gattung Spirogyra mit etwa 200 Arten, die weit verbreitet auftritt und durch ihre schraubig gewundene, spiralige Anordnung der Plastiden kaum zu verwechseln ist.
Als zweite große Gruppe treten die Vertreter der Chlorophyta auf.
Sie umfassen neben etlichen kleineren Gruppen, deren Einordnung umstritten ist, vor allem die Ulvophyceae und die Chlorophyceae.
Gerade die Chlorophyceae bilden nun eine weit verbreitete und artenreiche Gruppe.
Von den 4500 Arten der Chlorophyta entfallen 2300 auf die Chlorophyceae.
In dieser Gruppe finden wir die ganze Variationsbreite der Algen, von begeißelten Einzellern über alle Zwischenstufen bis hin zu koloniebildenden Arten
-
Volvocales (einschließlich Tetrasporales)
Einzellige oder koloniebildende Arten.
Die vegetative Vermehrung erfolgt durch Teilung innerhalb der Zellwand;
sobald die Teilung (eine oder auch mehrere) abgeschlossen ist, wird die alte, äußere Zellwand aktiv abgebaut, platzt auf und entläßt die Tochterzellen in die Umgebung.
Koloniebildende Arten erzeugen eine stets artspezifisch gleiche Anzahl von Zellen innerhalb der Kolonie; sie gehen stets auf eine Mutterzelle zurück.
Bekanntestes Beispiel dieser Gruppe ist Volvox.
Die kugelförmigen Kolonien von Volvox können mehrere tausend Einizeltiere umfassen und über einem Millimeter Durchmesser erreichen.
-
Chlorococcales

|
|
Pediastrum sp.
|
Sie zeichnen sich durch feste Zellwände und fehlende Geißeln aus.
Ebenso fehlen pulsierende Vakuolen.
Die Vermehrung erfolgt fast immer ungeschlechtlich.
Dabei wächst aus einer Tochterzelle innerhalb des elterlichen Verbands ein neuer Zell-Verband heran, der sich schließlich löst.
Bekannte Vertreter und leicht zu identifizieren sind die Pediastrum-Arten, die regelmäßig angeordnete Kolonien bilden.
Auch Scenedesmus ist leicht zu identifizieren.
Die Gattung Chlorella dient als Standardorganismus zur Erforschung der Photosynthese.
Auch das Wassernetz Hydrodictyon ist hier zu finden;
es bildet aus langen gestreckten Zellen, die an der Spitze verbunden bleiben ein dreidimensionales Geflecht von beachtlicher Größe.
-
Chaetophorales
Hier finden sich vor allem fädige Formen, die auf einer Oberfläche aufsitzen.
Die Fäden sind teils verzweigt, teils unverzweigt und wachsen an der Spitze.
Oft wächst der eine Teil kriechend am Substrat entlang, während aufragende Fäden die Reproduktionsorgane ins freie Wasser halten.
-
Oedogoniales
Fandenbildende Vielzeller; meist unverzweigt wachsend.
Ihr Charakteristikum bildet eine ungewöhnliche Art der Zellteilung, bei der Kernteilung und die Neubildung der Zellwand synchronisiert sind.
-
Kieselalgen

|
|
Kieselalge.
|
Charakteristisch für die formenreiche Gruppe der Kieselalgen (Diatomeen; Bacillariophyta) ist die Zellhülle aus Siliziumdioxid,
die sie aus im Wasser gelöster Kieselsäure synthetisieren und die diese Einzeller hart und widerstandsfähig macht.

|
|
Aufwuchs.
|
Kieselalgen treten in zwei Gruppen auf: Die zentrischen Kieselalgen (Centrales) treten als kreisrunde oder dreieckige Formen auf, die pennanten Varianten bilden langgestreckte stabförmige, mitunter gebogene oder oder S-förmige Formen.
Die pennanten Formen scheinen dabei bei weitem die häufigeren zu sein.
Manche Arten sitzen als Aufwuchs an kurzen Stilen mitunter massenhaft an Algenfäden.

|
|
Pennante K.
|
Diese Schale besteht aus zwei Hälften, wobei der Rand der oberen und der Rand der unteren Schale sich überlappen - ähnlich wie bei einer Pralinenschachtel.
Die obere Hälfte (Epitheka) ist daher stets etwas größer als die untere (Hypotheka).
Bei der Teilung wird stets eine Hypotheka gebildet, also die kleinere Hälfte.
Soweit die Vermehrung durch Teilung erfolgt ist also eine Tochterzelle stets etwas kleiner - im Laufe der Generationen verzwergen die Individuen.

|
|
Kieselalgen.
|
Ist eine Grenzgröße erreicht, setzt die sexuelle Phase ein.
Bei zentrischen Diatomeen entstehen Ei- und Spermazellen, die ins Wasser abgegeben werden, verschmelzen und als Auxospore zunächst ein Ruhestadium durchleben, bevor sie zu Originalgröße heranwachsen.
Pennante Kieselalgen betreiben Konjugation:
Dabei lagern sich zwei Diatomeen dicht aneinander und bilden eine gemeinsame Öffnung; ein Plamsakanal entsteht, über den Erbgut ausgetauscht wird.
Aus den neu variiertem Erbgut nach der wechselseitungen Verschmelzung der Zellkerne entstehen wiederum Auxosporen.

|
|
Kieselalgen.
|
Die beiden Schalen weisen eine phantastische Formenvielfalt auf; Linien- und Gitterstrukturen treten in großer Vielfalt auf und sind oft unerläßlich für die genaue Bestimmung der Art.
Dazu wird die Kieselalge von allem organischen Material befreit (Mazeration), etwa durch Aufkochen in Schwefelsäure, und bei 1000facher Vergrößerung untersucht.

|
|
Fortbewegung.
|
Mitunter können sich manche Arten von Kieselalgen aktiv fortbewegen (nicht die Centrales).
Dazu befindet sich auf der Körperunterseite in Längsrichtung ein Kanal (Raphe), der vorne, mittig und am Ende mit einer Pore ins Körperinnere versehen ist.
Zur Fortbewegung wird an der entsprechenden Pore eine klebrige Flüssigkeit abgesondert, die den Kanal entlangläuft und von der nächsten Pore aufgenommen wird.
Mit diesem raupenschlepperartigem Vortrieb kann eine Kieselalge in wenigen Sekunden seine eigene Körperlänge gleitend durchwandern.
(Im Mikroskop ein gespenstischer Anblick.)

|
|
Kieselalgen.
|
Kieselalgen haben global gesehen (zusammen mit den Dinoflagellaten) den größten Anteil am pflanzlichen Plankton und bilden mithin die wichtigste Basis der Nahrungskette.
Sie sind sowohl Hauptproduzent der organischen Stoffe als auch Haupterzeuger des atmosphärischen Sauerstoffs.
Die Ablagerungen ihrer Schalen (Kieselgur, Diatomeenerde) werden vielfältig genutzt, z.B. als Filtermaterial, Isolierung oder in Zahnpasta.
Sie treten von der Antarktis bis zur Arktis auf und sind auch in feuchten Böden zu finden.
Der braune Belag, der im zeitigen Frühjahr unter Wasser gelegene Steine überzieht sind vor allen Kieselalgen.
Ihre braune Färbung verdanken sie dem Pigment Fucoxanthin, der das Grün des Chlorophylls überdeckt.
-
Blaualgen
Sie zählen nur aus traditionellen Gründen zu den Algen, tatsächlich sind sie prokariotische Einzeller; im Grunde handelt es sich um stets unbegeißelte Bakterien.
Sie bilden entweder Kolonien, die von einer Gallertschicht umgeben sind oder fädige Formen, wobei auch der Faden von einer ausgeschiedenen Schleimschicht umgeben ist.
Blaualgen besitzen als Prokarioten keine Chloroplasten, stattdessen verfügen sie als photosynthetisch aktive Pigmente Chlorophyll a und Phycocyan, manchmal auch das rote Phycoerythrin, dazu auch Xanthophyll und Carotin.
Sie und das Mischungsverhältnis bestimmen die Färbung, die von gelbbraun über grün-blau bis fast schwarz variiert.
Blaualgen vermehren sich nur ungeschlechtlich durch Teilung oder Sporenbildung bei einzelligen oder koloniebildenden Arten, oder durch das Abtrennen von kurzen Fadenstücken bei fädigen Formen.
Manchmal erscheint es sinnvoller, Darwins Baum des Lebens zu ersetzen durch einen Fluß, der eine Schöpfungshöhe nicht kennt, sondern mit Gleichmut durch die Jahrmillionen fließt und die Form nimmt die ihm Täler und Kurven geben.
Dabei verzweigt und verästelt er sich immer wieder, manche Äste trocknen aus, andere erstarken mächtig und wieder andere Äste fließen wieder zusammen nachdem sie über Äonen getrennt waren.
Das daraus entstehende Gemenge so zu ordnen dass es am Ende aussieht wie das Dateisystem auf einer Festplatte ist ein Unterfangen, das wohl stets ein Versuch bleiben muss.
Da die Aufnahme eines externen photosynthetischen Organismus mehrfach geschehen ist finden sich bei den Algen daher zwangsläufig Vertreter verschiedener Großgruppen.
Grüalgen werden nach heutigem Stand (Adl u.a. 2005) zur Großgruppe der Archaeplastida gezählt.
Manche Arten von Euglena betreiben Photosynthese und werden daher zu den Algen gerechent. Sie stammen aus der Großgruppe der Excavata.
Braunalgen, Kieselalgen und Gelbgrüne Algen stammen aus der Großgruppe der Chromalveolata, sie haben ihren photosynthetisch aktiven Plastiden sekundär erworben.
(Von Wimpertierchen wird vermutet, sie könnten ursprünglich alle einen grünen Plastiden besessen und ihn wieder verloren haben.
Auch sie sind heute in die Chromalveolata eingeordnet.
Aus der gleiche Gruppe stammt der Einzeller Plasmodium falciparum, der Erreger der Malaria.
In ihm wurde ein geschrumpfter Plastid nachgewiesen, der von einer eukariotischen Alge stammt - der dauerhafte Verlust eines Plastiden scheint also nichts ungewöhnliches zu sein.
Malaria wird wohl tatsächlich von einer parasitisch gewordenen Alge verursacht.)
Geißeltierchen

|
|
Heterokonte Alge.
|
Geißeltierchen werden auch Flagellaten bezeichnet - Flagellum bezeichnet lateinisch eine Geißel oder Peitsche.
( Diejenigen Menschen, die sich aus religiösen Gründen selbst oder gegenseitig auspeitschen werden im Unterschied dazu Flagellanten genannt. Wer Monty Python kennt: Holzbretter tuns auch.)
Die namensgebende Geißel ist genauso aufgebaut wie eine Wimper der Wimpertierchen, meist jedoch deutlich länger und daher auffälliger.
Wimpern und Geißeln als ihre längere Variante sind sehr einheitliche aufgebaut und bestehen im Querschnitt stets aus neun kreisförmig angeordneten Doppelsträngen aus je zwei Mikrotubuli.
In der Mittelachse der Geißel verläuft ein weiterer Doppelstrang aus einem weiteren Paar Mikrotubuli.
Die Mikrotubuli sind röhrenförmige Strukturen aus Protein.
Unter Energieverbrauch verschieben sich die Mikrotubuli etwas gegeneinander und erzeugen so eine Krümmung der Geißel.
(Ein Kontraktion der Geißel ist nicht möglich.)
Bisweilen trägt die Geißel ihrerseits viele kleine Filamente (Mastigonemen oder Flimmern), die längs in regelmäßigen Abständen von der Geißel abzweigen.
Die Geißel erscheint fächer- oder federförmig.
Diese Flimmergeißeln erzeugen besonders viel Schub oder dienen einer speziellen Aufgabe.
Die Geißel kann am Hinterende Schub erzeugen oder auch am Vorderende des Körpers sitzen, wo sie mit ausgreifenden Bewegungen auf das umgebende Wasser eine Kraft ausübt, die das Geißeltierchen vorwärts zieht (Zuggeißel).
Auch seitlich aufsitzende Geißeln sind vereinzelt bekannt.
Wie viele andere Gruppen stellt auch die Gruppe der Geißeltierchen keine Verwandschaftsbeziehung dar.
Man sah unter dem Mikroskop eine Geißel und entschied sich bis auf seiteres, Geißelträger als Gruppe zusammenzufassen.
(So als ob man Erdmännchen, Menschen und Vögel in die Gruppe der Zweibeiner einordnet
und ihnen die Gruppe der Vierbeiner gegenüberstellt.
Kletteraffen, die sich mit ihrem Schwanz durch die Bäume hangeln bilden dann die Gruppe der Fünfbeiner, allerdings incertae sedis, also nicht sicher.)
Innerhalb der Gruppe lassen sich pflanzliche Flagellaten, die Photosynthese betreiben, von tierischen Flagellaten unterscheiden, die sich von fremden organischen Substanzen ernähren.
(Phytoflagellaten / Zooflagellaten).

|
|
Zuggeißel.
|
Auch viele Bakterien besitzen einen fadenförmigen Fortsatz, der zur Fortbewegung dient.
Da er aber gänzlich anders aufgebaut ist als die Geißel der eukariotischen Zellen und nicht schlägt sondern rotiert, wird er Flagellum genannt, auch wenn diese Unterscheidung nicht immer durchgehalten wird.
(Auch dieser völlig unterschiedliche Bau ist ein weiterer guter Grund, die Organismen in Eukarioten und Prokarioten zu unterteilen.)
Die Fähigkeit, Geißeln hervorzubringen ist auch bei Säugetieren nicht ganz verloren gegangen, das Unterprogramm in den Tiefen der DNS kann immer noch abgerufen werden.
Die Oberfläche der Luftröhre sind mit Geißeln ausgestattet, die Staub wieder in Richtung Luftröhreneingang zurücktransportieren, wo er ausgehustet werden kann;
auch Spermazellen stellen begeißelte Zellen dar.
Geißeltierchen sind oft sehr schnell und klein.
Unter dm Mikroskop sind sie schwer zu verfolgen, ihre geringe Körpergröße macht es ihnen möglich,
auch bei einem relativ flach aufliegendem Deckglas zwischen oberem und unterem Ende der Wasserschicht hin und her zu schwimmen, so dass sie flugs aus dem Schärfebereich herauswandern.
Sie sind die scheuen Geparden der Serengeti.
Bauchhärlinge

|
|
Bauchhärling.
|
Bauchhärlinge (Gastrotricha) sind kleine mehrzellige Tiere, die im Süßwasser und im Meer vorkommen.
Allen gemeinsam ist ein in etwa flaschenartiger Körperbau ( der Leibesumfang ist etwa ab der Körpermitte verdickt)
und paarig auftretende, stachelförmige Ausläufer am Körperende.
Die flache Bauchseite ist dicht besetzt mit Flimmerhärchen; ihre Bewegungen schieben den Bauchhärling gleitend vorwärts.
Gemeinsam mit den Nematoden ist ihnen das Fehlen von Ringmuskulatur
(Muskelkontraktionen längs der Körperachse werden mit dem Körperinnendruck wieder ausgeglichen),
der Saug- und Pressmagen und die Körperaußenhaut, in der die Zellwände aufgelöst sind
und das Gewebe aus Zellmaterial und vielen Zellkernen besteht (Synzytium).
Bauchhärlinge ernähren sich von Bakterien, Algen und Detritus.
Fortpflanzung erfolgt vielfältig: Je nach Art kommen Jungfernzeugung, Paarungen oder Zwitter mit Selbstbefruchtung vor.
Aus den Eiern schlüpfen nach ca. 2 Tagen Jungtiere, ein Larvenstadium existiert nicht.
Die Einordnung ist unklar - gleichen sie äußerlich den Nematoden und Rotatorien, so stehen sie genetisch den Plattwürmern näher.
Nematoden

|
|
Nematode.
|
Nematoden sind kleine vielzellige Tiere, die eine überaus artenreiche Gruppe bilden.
Sie treten stets als fadenförmige, langgestreckte dünne wurmartige Tiere in Erscheinung, daher werden sie auch als Fadenwürmer bezeichnet.
Da sie ihren Körperbau abgesehen von der Größe kaum abwandeln ist eine Unterscheidung der Arten oft kaum möglich.
Meist leben sie in feuchtem Boden aber auch im Süßwasser und im Meer.
Da die Nematoden durch Häutung wachsen sind sie näher mit den Insekten oder Krebstieren verwandt und nicht mit den bekannten Ringelwürmern (von denen der Regenwurm aus dem Gartenboden der bekannteste sein dürfte).
Der Körper eines Nematoden ist nicht gegliedert, sondern besteht aus einer einheitlichen langen Röhre, die an Kopf und Hinterende ausgezogen ist.
Der Kopf beginnt mit der zentralen Mundöffnung, an der je nach Art einige Fortsätze der Nahrungsaufnahme dienen.
Da sich die zahlreichen Nematodenarten sehr unterschiedlich ernähren ist der Mundapparat bei genauerem Hinsehen entsprechend vielgestaltig.
Nematoden sind oft Detritusfresser, Bakterienfresser oder saugen an Mulm und abgestorbenen Pflanzenteilen;
andere erbeuten Kieselalgen, Fadenalgen oder auch kleinere Beutetiere wie kleine Rotatorien.
Wieder andere stechen pflanzliche Zellen an und saugen sie aus.
Nematoden verfügen über kein Zirkulationssysten wie einen Blut- oder Körperflüssigkeitskreislauf.
Sie besitzen keine Atmungsorgane und nur ein sehr schlichtes Nervensystem.
Nematoden sind zellkonstant.

|
|
N. im Dickicht.
|
Sie verfügen praktisch nur über Längsmuskeln, die den Körper in Längsrichtung kontrahieren.
Ringmuskeln fehlen.
Als Gegenspieler fungiert der Leibesinnendruck, der von einer stabilen, widerstandsfähigen Haut aufrecht erhalten wird.
Nach einer Kontraktion einer Körperseite, die den Nematoden in einer Richtung krümmt, ist es der Innendruck, der bei nachlassender Muskelkraft den Körper wieder gerade biegt;
im Anschluss werden die Muskeln der gegenüberliegenden Seite kontrahiert und das Tier krümmt sich in die andere Richtung.
In schnellen Wechselspiel wird eine schlängelnde Bewegung erzeugt, die für den Vortrieb sorgt.
Die beiden Muskelstränge liegen dabei bauchseitig und rückseitig, die Schlängelbewegung läuft also auf und ab.
Wenn unter der Lupe ein Nematode seitwärts zu schlagen scheint, liegt er in Wirklichkeit in Seitenlage und schlängelt nach wie vor auf und ab.
Nematoden pflanzen sich stets getrenntgeschlechtlich fort.
Männchen besitzen am Hinterende paarige ausfahrbare Fortsätze, mit deren Hilfe sie die Weibchen beim Paarungsakt positionieren.
Weibchen - sofern sie durchsichtig sind - erkennt man an den Embryos in ihrem Körperinneren.
Diese Geschlechtsmerkmale sind bei Nematoden wie dem Mikrowürmchen (Panagrellus redivivus) sehr gut sichtbar, bei Fadenwürmern die unter Wasser leben habe ich sie jedoch nie erkennen können.
Auch zappeln die untergetaucht lebenden Nematoden unablässig und beharrlich, so dass sie sich einer Beobachtung hartnäckig widersetzen und auch das umgebende Medium derart in Unruhe versetzen, dasss ihre Anwesenheit oftmals einer Geduldsprobe gleichkommt.
Links und Buchempfehlungen
***
-
Streble / Krauter: Das Leben im Wassertropfen. Kosmos Verlag. Das Standardwerk schlechthin mit hervorradenden Zeichnungen und Fotos zur Bestimmung zahlreiche Einzeller und kleiner Vielzeller im gemäßigten Süßwasser.Mittlerweile in der 10 + x ten Auflage.
-
Linne von Berg, Hoef-Emden, Marin, Melkonian: Der Kosmos-Algenführer. 2012, Kosmos Verlag. Durchdachter Bestimmungschlüssel und zahlreiche ausgezeichnete Fotos ermöglichen die Bestimmung von über 150 Arten von heimischen Süßwasseralgen.
-
Wilhelm Foissner, Fritz Kohmann: Bestimmung und Ökologie der Mikrosaprobien nach DIN 38410. Gustav Fischer Verlag.
-
Max Mayer: Kultur und Präparation der Protozoen. Kosmos-Verlag. Frankhsche Verlagshandlung. Stuttgart 1981.
letzte Änderung: 25. Mai 2014
|
|
|
.: 
 :.
:.
|
|























")



")
























